脑未来科普 | 神经活动的行进波
|
|
电活动-具体表现为神经元膜电位的去极化-是单个神经元活动的一般表现形式。而在由大量相互连接的神经元组成的神经网络中,这些神经元的电活动往往不是随机无规律的,而是呈现出某种时空特征。这种时空特征的一种表现形式是呈现出在神经网络中波动推进的时空形态。这种波动推进的时空特征被称为神经活动的行进波(traveling wave)。
|

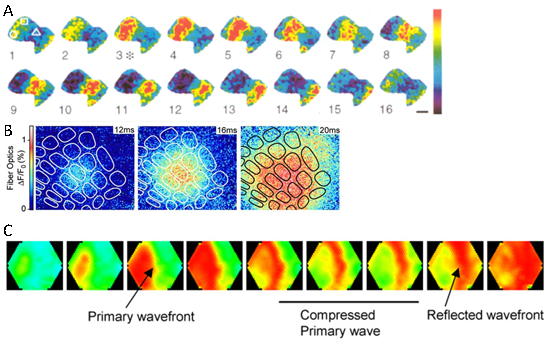
图1.不同VSD成像实验中观察到的神经活动的行进波 A.大蛞蝓的嗅觉前脑叶,引自参考资料[2];B.清醒小鼠的胡须感觉皮层,引自参考资料[3];C.麻醉活体大鼠的初级视觉皮层和高级视觉皮层,引自参考资料[4]。
科学家们一直在试图探究这种行进波的生理意义。同时,科学家们也不确定这种行进波是否在活的动物的神经系统同样存在。于是,首先人们在活的麻醉动物上,如龟的视觉皮层上以及雪貂和猫的大脑皮层都观察到了这种行进波的存在;同时,研究者们也在清醒的大鼠和猴子的大脑皮层观察到了这种去极化波的存在[3, 5]。进入新世纪之后,人们对于这种行进波的作用的研究越来越多。研究发现,这种去极化波可能在脑区内部起着信息交互的作用。比如,在啮齿类动物,如大鼠和小鼠中,胡须被认为是比眼睛还要重要的感觉器官,因此大鼠和小鼠的胡须感觉皮层上专门形成了很多的圆柱状小区域(英文称为barrel小区),每个barrel小区特异地对动物的一根胡须的触觉信号有反应。当这根胡须被触动时,相应的barrel小区被兴奋,接着,这种兴奋通过行进波的形式传导到周边的barrel小区[3](图1.B)。这样,当有多根胡须同时被触动时(这在实际动物生活场景中常常会出现),这种神经兴奋的行进波就可以帮助整合多个barrel小区的兴奋信息。这种行进波的速度被认为与皮层表层的神经元轴突的非髓鞘化传输速度有关[6]。类似的,这种神经兴奋的行进波还可以在不同脑区之间传递信息,比如在初级视觉皮层和高级视觉皮层之间传递视觉信息[4](图1.C),以及在身体感觉皮层和运动皮层之间整合感觉和运动信息[7]。
以上这些是在动物实验中观测到的结果,那么,在人类被试大脑中是否也能观测到这种神经兴奋的行进波呢?答案是有的。在脑电图(electroencephalogram, EEG)的脑电记录中(EEG记录电极阵列呈二维分布,可以记录电活动的时空特征),可以清楚地观测到特定频段脑电波活动的时空分布呈现行进波的特征[8](图2.A)。与EEG记录类似的,在脑皮层电图(electrocorticography, ECoG)的脑电记录中,也可以发现皮层的特定频段的电活动呈现从颞叶到顶叶再到前额叶的行进波传播特征[9](图2.B)。这种全脑范围的神经活动行进波的速度可能与脑区之间大脑白质的髓鞘化传输速度有关[6]。而这种行进波的功能则可能与不同脑区之间的功能连接,以及睡眠过程中的记忆巩固[9]有关。
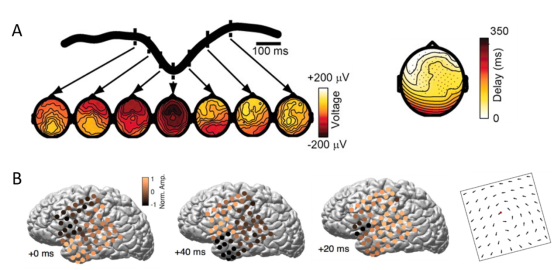
图2.EEG(图A,引自参考资料[8])和ECoG(图B,引自参考资料[9])记录的脑电数据中的神经活动的行进波。
那么这种神经系统中神经活动行进波的形成机制是什么呢?对此,研究者们有两种理论猜测:1.波的运动来自一个共同的振荡子,这个振荡子直接把兴奋传递到相邻的皮层脑区,因为这种兴奋传递到不同脑区有时间差,所以形成了这些脑区活动之间的相位差(图3.A);2.波的运动起始于一个单一的振荡子,这个振荡子把其自身的周期性发放脉冲沿皮层逐区域传播,这一传播依赖于皮层各区域之间的连接(图3.B)[10]。
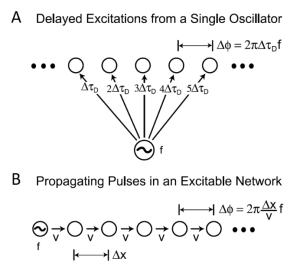
图3.两种可能的神经活动行进波的形成机制,引自参考资料[10]。
综上,神经系统中神经活动行进波是神经网络中神经细胞群体电活动所表现出来的时空特征。其生理功能可能是神经系统各子区域之间互相作用和相互联系的重要形式。无论是光学成像记录结果还是脑电记录结果都验证了这种神经活动行进波的存在。未来,随着科学家的深入研究,我们有望揭示神经活动行进波的功能和机制。
参考文献
1. Grinvald, A., A. Manker, and M. Segal, Visualization of the spread of electrical activity in rat hippocampal slices by voltage-sensitive optical probes. The Journal of Physiology, 1982. 333(1): p. 269-291.
Abstract: 1. Voltage-sensitive membrane-bound dyes and a matrix of 100 photodetectors were used to detect the spread of evoked electrical activity at the CA1 region of rat hippocampus slices. A display processor was designed in order to visualize the spread of electrical activity in slow motion.2. The stimulation of the Schaffer collateral-commissural path in the stratum radiatum evoked short latency (2-4 msec) fast optical signals, followed by longer latency (4-15 msec) slow signals which decayed within 20-50 msec. Multiple fast signals were frequently detected at the stratum pyramidale; they propagated toward the stratum oriens with an approximate conduction velocity of 0.1 m/sec.3. The fast signals were unaltered in a low Ca2+ high Mg2+ medium but were blocked by tetrodotoxin. These signals probably represent action potentials in the Schaffer collateral axons. Their conduction velocity was about 0.2 m/sec and their refractory period about 3-4 msec.4. The slow signals were absent in a low Ca2+ medium and probably represent excitatory post-synaptic potentials (e.p.s.p.s) generated in the apical dendrites of the pyramidal cells. They were generated in the stratum radiatum, where the presynaptic signals were seen, and spread into somata and basal dendrites (the stratum pyramidale and oriens, respectively).5. The timing of the signals with fast rise-time, which were detected at the statum pyramidale, approximately coincided with the timing of the extracellularly recorded field potentials. These multiple discharges probably represent action potentials of the pyramidal cells. They spread back into the apical dendrites but with significant attenuation of the amplitudes of the high frequency components of the pyramidal action potentials.6. Hyperpolarizing potentials could be detected when strong stimuli were applied to the stratum radiatum or alveus. The net hyperpolarizations were detected only in the stratum pyramidale and the border region between the stratum pyramidale and radiatum. Frequently the inhibition was masked by the large e.p.s.p.s. However, its existence could be demonstrated by treatment of the slice with picrotoxin or a low Cl- medium. Under these conditions a long-lasting depolarization of the apical dedrites was evoked by the stimulation. This was associated with an increase of the multiple discharges in the stratum pyramidale and oriens.7. These studies illustrate the usefulness of voltage-sensitive dyes in the analysis of passive and active electrical properties, pharmacological properties and synaptic connexions in mammalian brain slices, at the level both of small neuronal elements (dendrites, axons) and of synchronously active neuronal populations.
2. Kleinfeld, D., et al., Dynamics of propagating waves in the olfactory network of a terrestrial mollusk: an electrical and optical study. J Neurophysiol, 1994. 72(3): p. 1402-1419.
Abstract: 1. The procerebral (PC) lobe of the terrestrial mollusk Limax maximus contains a highly interconnected network of local olfactory interneurons that receives ipsilateral axonal projections from superior and inferior noses. This network exhibits an approximately 0.7-Hz intrinsic oscillation in its local field potential (LFP). 2. Intracellular recordings show that the lobe contains at least two classes of neurons with activity phase locked to the oscillation. Neurons in one class produce periodic bursts of spikes, followed by a period of hyperpolarization and subsequently a depolarizing afterpotential. There is a small but significant chance for a second burst to occur during the depolarizing afterpotential; this leads to a double event in the LFP. Bursting neurons constitute approximately 10% of the neurons in the lobe. 3. Neurons in the other class fire infrequently and do not produce periodic bursts of action potentials. However, they receive strong, periodic inhibitory input during every event in the LFP. These nonbursting cells constitute the major fraction of neurons in the lobe. There is a clear correlation between the periodic burst of action potentials in the bursting neurons and the hyperpolarization seen in nonbursting neurons. 4. Optical techniques are used to image the spatially averaged transmembrane potentials in preparations stained with voltage-sensitive dyes. The results of simultaneous optical and electrical measurements show that the major part of the optical signal can be interpreted as a superposition of the intracellular signals arising from the bursting and nonbursting neurons. 5. Successive images of the entire PC lobe show waves of electrical activity that span the width of the lobe and travel its full length along a longitudinal axis. The direction of propagation in the unperturbed lobe is always from the distal to the proximal end. The wavelength varies between preparations but is on the order of the length of the preparation. 6. One-dimensional images along the longitudinal axis of the lobe are used to construct a space-time map of the optical activity, from which we calculate the absolute contribution of bursting and nonbursting neurons to the optical signal. The contribution of the intracellular signals from the two cell types appears to vary systematically across the lobe; bursting cells dominate at middle and proximal locations, and nonbursting cells dominate at distal locations. 7. The direction and form of the waves can be perturbed either by microsurgical manipulation of the preparation or by chemical modulation of its synaptic and neuronal properties.
3. Ferezou, I., S. Bolea, and C.C. Petersen, Visualizing the cortical representation of whisker touch: voltage-sensitive dye imaging in freely moving mice. Neuron, 2006. 50(4): p. 617-29.
Abstract: Voltage-sensitive dye imaging resolves the spatiotemporal dynamics of supragranular subthreshold cortical activity with millisecond temporal resolution and subcolumnar spatial resolution. We used a flexible fiber optic image bundle to visualize voltage-sensitive dye dynamics in the barrel cortex of freely moving mice while simultaneously filming whisker-related behavior to generate two movies matched frame-by-frame with a temporal resolution of up to 2 ms. Sensory responses evoked by passive whisker stimulation lasted longer and spread further across the barrel cortex in awake mice compared to anesthetized mice. Passively evoked sensory responses were large during behaviorally quiet periods and small during active whisking. However, as an exploring mouse approached an object while whisking, large-amplitude, propagating cortical sensory activity was evoked by active whisker-touch. These experiments demonstrate that fiber optics can be used to image cortical sensory activity with high resolution in freely moving animals. The results demonstrate differential processing of sensory input depending upon behavior.
4. Xu, W., et al., Compression and reflection of visually evoked cortical waves. Neuron, 2007. 55(1): p. 119-29.
Abstract: Neuronal interactions between primary and secondary visual cortical areas are important for visual processing, but the spatiotemporal patterns of the interaction are not well understood. We used voltage-sensitive dye imaging to visualize neuronal activity in rat visual cortex and found visually evoked waves propagating from V1 to other visual areas. A primary wave originated in the monocular area of V1 and was "compressed" when propagating to V2. A reflected wave initiated after compression and propagated backward into V1. The compression occurred at the V1/V2 border, and local GABAA inhibition is important for the compression. The compression/reflection pattern provides a two-phase modulation: V1 is first depolarized by the primary wave, and then V1 and V2 are simultaneously depolarized by the reflected and primary waves, respectively. The compression/reflection pattern only occurred for evoked waves and not for spontaneous waves, suggesting that it is organized by an internal mechanism associated with visual processing.
5. Slovin, H., et al., Long-Term Voltage-Sensitive Dye Imaging Reveals Cortical Dynamics in Behaving Monkeys. Journal of Neurophysiology, 2002. 88(6): p. 3421-3438.
Abstract: A novel method of chronic optical imaging based on new voltage-sensitive dyes (VSDs) was developed to facilitate the explorations of the spatial and temporal patterns underlying higher cognitive functions in the neocortex of behaving monkeys. Using this system, we were able to explore cortical dynamics, with high spatial and temporal resolution, over period of ≤1 yr from the same patch of cortex. The visual cortices of trained macaques were stained one to three times a week, and immediately after each staining session, the monkey started to perform the behavioral task, while the primary and secondary visual areas (V1 and V2) were imaged with a fast optical imaging system. Long-term repeated VSD imaging (VSDI) from the same cortical area did not disrupt the normal cortical architecture as confirmed repeatedly by optical imaging based on intrinsic signals. The spatial patterns of functional maps obtained by VSDI were essentially identical to those obtained from the same patch of cortex by imaging based on intrinsic signals. On comparing the relative amplitudes of the evoked signals and differential map obtained using these two different imaging methodologies, we found that VSDI emphasizes subthreshold activity more than imaging based on intrinsic signals, that emphasized more spiking activity. The latency of the VSD-evoked response in V1 ranged from 46 to 68 ms in the different monkeys. The amplitude of the V2 response was only 20−60% of that in V1. As expected from the anatomy, the retinotopic responses to local visual stimuli spread laterally across the cortical surface at a spreading velocity of 0.15−0.19 m/s over a larger area than that expected by the classical magnification factor, reaching its maximal anisotropic spatial extent within ∼40 ms. We correlated the observed dynamics of cortical activation patterns with the monkey's saccadic eye movements and found that due to the slow offset of the cortical response relative to its onset, there was a short period of simultaneous activation of two distinct patches of cortex following a saccade to the visual stimulus. We also found that a saccade to a small stimulus was followed by direct transient activation of a cortical region in areas of V1 and V2, located retinotopically within the saccadic trajectory.
6. Muller, L., et al., Cortical travelling waves: mechanisms and computational principles. Nature Reviews Neuroscience, 2018. 19(5): p. 255-268.
Abstract: Multichannel recording technologies have revealed travelling waves of neural activity in multiple sensory, motor and cognitive systems. These waves can be spontaneously generated by recurrent circuits or evoked by external stimuli. They travel along brain networks at multiple scales, transiently modulating spiking and excitability as they pass. Here, we review recent experimental findings that have found evidence for travelling waves at single-area (mesoscopic) and whole-brain (macroscopic) scales. We place these findings in the context of the current theoretical understanding of wave generation and propagation in recurrent networks. During the large low-frequency rhythms of sleep or the relatively desynchronized state of the awake cortex, travelling waves may serve a variety of functions, from long-term memory consolidation to processing of dynamic visual stimuli. We explore new avenues for experimental and computational understanding of the role of spatiotemporal activity patterns in the cortex.
7. Ferezou, I., et al., Spatiotemporal dynamics of cortical sensorimotor integration in behaving mice. Neuron, 2007. 56(5): p. 907-23.
Abstract: Tactile information is actively acquired and processed in the brain through concerted interactions between movement and sensation. Somatosensory input is often the result of self-generated movement during the active touch of objects, and conversely, sensory information is used to refine motor control. There must therefore be important interactions between sensory and motor pathways, which we chose to investigate in the mouse whisker sensorimotor system. Voltage-sensitive dye was applied to the neocortex of mice to directly image the membrane potential dynamics of sensorimotor cortex with subcolumnar spatial resolution and millisecond temporal precision. Single brief whisker deflections evoked highly distributed depolarizing cortical sensory responses, which began in the primary somatosensory barrel cortex and subsequently excited the whisker motor cortex. The spread of sensory information to motor cortex was dynamically regulated by behavior and correlated with the generation of sensory-evoked whisker movement. Sensory processing in motor cortex may therefore contribute significantly to active tactile sensory perception.
8. Massimini, M., et al., The Sleep Slow Oscillation as a Traveling Wave. The Journal of Neuroscience, 2004. 24(31): p. 6862-6870.
Abstract: During much of sleep, virtually all cortical neurons undergo a slow oscillation (<1 Hz) in membrane potential, cycling from a hyperpolarized state of silence to a depolarized state of intense firing. This slow oscillation is the fundamental cellular phenomenon that organizes other sleep rhythms such as spindles and slow waves. Using high-density electroencephalogram recordings in humans, we show here that each cycle of the slow oscillation is a traveling wave. Each wave originates at a definite site and travels over the scalp at an estimated speed of 1.2-7.0 m/sec. Waves originate more frequently in prefrontal-orbitofrontal regions and propagate in an anteroposterior direction. Their rate of occurrence increases progressively reaching almost once per second as sleep deepens. The pattern of origin and propagation of sleep slow oscillations is reproducible across nights and subjects and provides a blueprint of cortical excitability and connectivity. The orderly propagation of correlated activity along connected pathways may play a role in spike timing-dependent synaptic plasticity during sleep.
9. Muller, L., et al., Rotating waves during human sleep spindles organize global patterns of activity that repeat precisely through the night. eLife, 2016. 5: p. e17267.
Abstract: During sleep, the thalamus generates a characteristic pattern of transient, 11-15 Hz sleep spindle oscillations, which synchronize the cortex through large-scale thalamocortical loops. Spindles have been increasingly demonstrated to be critical for sleep-dependent consolidation of memory, but the specific neural mechanism for this process remains unclear. We show here that cortical spindles are spatiotemporally organized into circular wave-like patterns, organizing neuronal activity over tens of milliseconds, within the timescale for storing memories in large-scale networks across the cortex via spike-time dependent plasticity. These circular patterns repeat over hours of sleep with millisecond temporal precision, allowing reinforcement of the activity patterns through hundreds of reverberations. These results provide a novel mechanistic account for how global sleep oscillations and synaptic plasticity could strengthen networks distributed across the cortex to store coherent and integrated memories.
10. Prechtl, J.C., T.H. Bullock, and D. Kleinfeld, Direct evidence for local oscillatory current sources and intracortical phase gradients in turtle visual cortex. Proc Natl Acad Sci U S A, 2000. 97(2): p. 877-82.
Abstract: Visual stimuli induce oscillations in the membrane potential of neurons in cortices of several species. In turtle, these oscillations take the form of linear and circular traveling waves. Such waves may be a consequence of a pacemaker that emits periodic pulses of excitation that propagate across a network of excitable neuronal tissue or may result from continuous and possibly reconfigurable phase shifts along a network with multiple weakly coupled neuronal oscillators. As a means to resolve the origin of wave propagation in turtle visual cortex, we performed simultaneous measurements of the local field potential at a series of depths throughout this cortex. Measurements along a single radial penetration revealed the presence of broadband current sources, with a center frequency near 20 Hz (gamma band), that were activated by visual stimulation. The spectral coherence between sources at two well-separated loci along a rostral-caudal axis revealed the presence of systematic timing differences between localized cortical oscillators. These multiple oscillating current sources and their timing differences in a tangential plane are interpreted as the neuronal activity that underlies the wave motion revealed in previous imaging studies. The present data provide direct evidence for the inference from imaging of bidirectional wave motion that the stimulus-induced electrical waves in turtle visual cortex correspond to phase shifts in a network of coupled neuronal oscillators.
—— 作者介绍 ——
 |
王志杰 2012年中国科学院生物物理所博士毕业,现为中国科学院深圳先进技术研究院脑认知与脑疾病研究所王立平课题组助理研究员。 研究方向:小鼠大脑皮层在情绪处理中的作用 |